04/07/2021
0
Изгледи

Митохондриалният геном е малка, затворена кръгова молекула с двойна спирала от 16 569 базови двойки (bp), която се наследява само от майки на синове и дъщери (рядко). случаи на хетероплазмия), така че няма генетична рекомбинация по време на зачеването, то също се развива изключително чрез натрупване на мутации в метеорологично време.





Високата скорост на мутация на митохондриалния геном (10 до 20 пъти по-висока от тази на ДНК ядрен, по отношение на гени със сравними функции) е полезен за разграничаване между популациите, които с течение на времето са били биологично свързани, но тяхната скорост на мутация може да бъде толкова бърза, че да възникнат мутационни явления. обратна мутация. Скоростта на еволюция Средната стойност на тази молекула е изчислена от видове, при които са налични времена на отклонение от вкаменелости, и от биодемографски или протеинови данни; резултатът дава 1-2% на милион години, валиден за различни поръчки.
Тази конкретна ДНК функционира като инструмент за изследвания биологичното родство между популациите, защото техните свойства позволяват да се изяснят връзките между популациите които са се разминавали в последно време, без да се налага да се вземат предвид явленията на рекомбинация и добавяне на измерение временно; За това трябва да се има предвид следното: два индивида, чийто общ прародител е жена, ще имат митохондриални ДНК молекули толкова различни, колкото времето е изминало от отделянето на прародител.
Някои региони на ДНК могат да бъдат много подобни един на друг, което позволява тяхното класификация в същия клъстер, който е известен като хаплогрупа. Например, Torroni et al (1993) описват основополагащите митохондриални линии за американския континент, които са получили наименованието на хаплогрупи A, B, C и D, според точкови мутации на последователността митохондриална. Тези мутации създават различни места на разцепване за специфични ензими, както е описано по-долу: в линия А има мястото прекъсване 663 за ензима HaeIII, линия C се характеризира с място 13259 за HincII, в линия D сайт 5176 се разпознава за АлуИ; тази идентификация се прави от полиморфизъм на дължината на фрагмента (RFLP). В случая на линия B, има заличаване на 9 базови двойки на позиция 8272-8289.
Географското разпределение на всяка линия е описано, както следва: линия А е преобладаваща в американския континент и особено в Северна Америка; но тази хаплогрупа се приписва и на мезоамериканските популации. Линии C и D се появяват главно в Южна Америка; линия B се намира в северния и южния регион на тихоокеанското крайбрежие и Kemp et al (2010) я предлага като характерна линия на семейство Юто-ацтека и югозападна част на САЩ.
Освен това има мутации, които се споделят между индивиди от една и съща хаплогрупа, което прави възможно описанието на специфични хаплотипове (или подлинии) на някои популации. Фигура 1 показва регионите на митохондриалната ДНК, които съдържат мутациите за разпознаване на всяка линия.
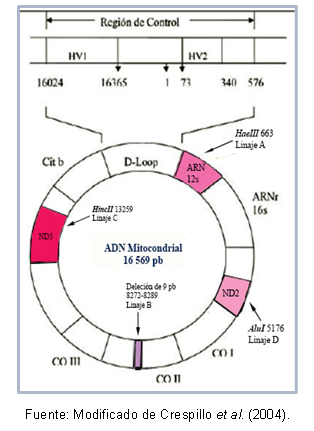
Фигура 1. Митохондриални ДНК региони, типични за всяка американска основополагаща линия.
В продължение на няколко години само тези четири линии бяха признати, докато хаплогрупа X не беше включена за северните популации на континента; което също е далечно свързано с европейските популации. Предишните линии имат съответствие с азиатските популации, въпреки че се наблюдават по-рядко, отколкото в американския континент; линии A, B и C не се срещат при съвременните африканци и кавказци; и линия D също съществува в Африка, но свързана с други рестрикционни сайтове. С горното можем да кажем, че тези хаплогрупи характеризират американските популации и следователно тяхното изследване е адекватно по отношение на заселването и миграциите (стари и скорошни).
Начинът за установяване на генетични прилики или разлики между популациите е с генетични разстояния, които могат да имат историческо обяснение, тъй като те се променят (увеличават или намаляват) през поколенията и могат да ни доведат до събития в историята от един население например: по време на голяма миграция или контакт между две култури, с това изчисление можем да различим какви механизми са се задействали, за да ни дадат този или онзи резултат.
Както генетичният дрейф, така и генният поток са свързани с лекотата, с която група, в този случай човешки същества, трябва да се движи в рамките на група. територия и да влизат в контакт с други групи. И така, географската изолация е генетичното разстояние, което расте с увеличаване на географското разстояние между човешките групи.
За да обясня разнообразие в основополагащите митохондриални линии в Америка има две хипотеза: че този континент е бил колонизиран от множество събития от Берингия или че след като е настъпила миграция, еволюционни промени са настъпили след колонизацията. Два пътя на навлизане на тези вариации на континента също са обяснени, като първият предполага, че четирите основополагащи хаплогрупи без вариации, тоест всяка с коренен хаплотип, те може да са пристигнали точно след последния ледников максимум или малко преди това, с дати отпреди 21 хиляди до 19 хиляди години и биха следвали крайбрежен маршрут от Спокоен; второто предложение предполага, че тези вътрешно-хаплогрупови вариации вече са съществували в Берингия и са били пренесени на юг от американския континент, но тяхното навлизане би било точно в края на последния ледников максимум, така че пътищата в рамките на континента вече да са свободни, тогава навлизането на тези човешки групи ще бъде преди 19 хиляди години. Освен това има голямо разнообразие от хаплогрупа А и време на сливане за нея, което е по-кратко от останалите (17 хиляди години).
Най-вероятното обяснение за това е, че се дължи на вторични разширения на хаплогрупа А от Берингия, дълго след края на последния ледников максимум. Въпреки несъответствието по отношение на времето на навлизане на човека в Америка, генетичните изследвания имат дават известна яснота, защото подкрепят хипотезата, че преди е имало човешки групи на американския континент Кловис; и се установява, че има разделение между предците на Североизточна Азия преди 25-35 хиляди години и навлизането в Америка преди 15-35 хиляди години. Фигура 2 показва маршрутите на митохондриалните хаплогрупи в света и времето на разминаване в години преди настоящето.
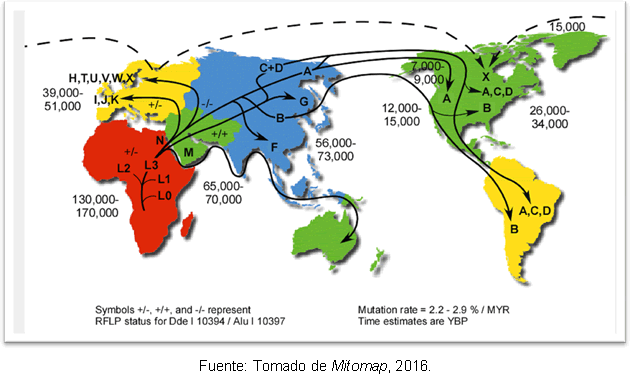
Фигура 2. Карта на различните пътища на разпространение на митохондриални линии
Има много полезен инструмент за генетичен анализ; филогеографията. Това са първите приложения на молекулярните изследвания и с тях се стремим да можем да определим филогенетичните и пространствени връзки между нуклеотидни последователности, в този случай ДНК митохондриална. Пространственото разпределение може да наподобява времеви модел, тоест най-географски отдалечените ДНК последователности трябва да са най-много генетично различни, както и ДНК последователности, които са се разминавали отдавна, също трябва да са най-различни генетично. Така че географски отдалечени популации, с малък или никакъв генен поток между тях, биха натрупали различия поради генетичен дрейф и мутация, дори чрез селекция; но могат да възникнат ситуации, които не позволяват дрейф, като един или повече основателни ефекти или други модели на генния поток.
Данните, предоставени от митохондриалните гени, са сред най-полезните във филогеографските изследвания поради характеристика на тези гени да не се рекомбинират и в резултат те показват много по-ясна филогенетична линия от много ядрени гени. Ефективният размер на популацията, изчислен с митохондриалните гени (и с Y хромозомата, функционираща по подобен начин) е около една четвърт от този, изчислен за митохондриалните гени. ядрени гени, тогава дивергенцията настъпва почти четири пъти по-бързо, отколкото при ядрените гени, тази бърза скорост на дивергенция (и генен поток) може да причини наблюдаваният модел на наследяване в тези еднородителски гени е различен от филогениите, получени с ядрени гени (които представляват по-голямата част от генофонда в индивидуален).
Кавали-Сфорца, Л. Л. (2000). Гени, народи и езици (1-во издание в изд. Pocket Library). Барселона, Испания: Критична редакция.
Rebato, E., Susanne, C., & Chiarelli, B. (Редакции.). (2005). Да разберем биологичната антропология: еволюцията и човешката биология. Божествено Слово.
Торони, А., Шур, Т. Г., Кабел, М. Ф, Браун, М. Д, Нийл, Дж. V., Larsen, M., et al. (1993). Азиатски афинитети и континентална радиация на четирите основополагащи индиански мтДНК. Американският журнал за човешка генетика, 53 (3), 563.
(2022). митохондриална ДНК, Национален институт за изследване на човешкия геном.
-Схематична справка за митохондриална ДНК-
Crespillo, M., Paredes, M., Arimany, J., Guerrero, L., & Valverde, J. (2004). Гражданска война в Испания (1936-1939): идентифициране на човешки останки от масови гробове в Каталуния чрез анализ на митохондриална ДНК. Относно случай. Тетрадки по съдебна медицина (38), 37-46.
