04/07/2021
0
Visninger

Det mitokondrielle genom er et lille, lukket cirkulært molekyle med dobbelt helix på 16.569 basepar (bp), der kun nedarves fra mødre til sønner og døtre (sjældent). tilfælde af heteroplasmi), så der er ingen genetisk rekombination på tidspunktet for undfangelsen, den udvikler sig også udelukkende ved akkumulering af mutationer i vejr.





Den høje mutationshastighed i mitokondriegenomet (10 til 20 gange så stor som DNA nuklear, hvad angår gener med sammenlignelige funktioner) er nyttig til at differentiere mellem populationer, der over tid har været biologisk beslægtede, men deres mutationshastighed kan være så høj, at mutationsfænomener kan forekomme. tilbagemutation. Satsen på udvikling Middelværdien af dette molekyle er blevet beregnet ud fra arter, hvor afvigelsestider fra fossile rester var tilgængelige, og fra biodemografiske eller proteindata; resultatet giver 1-2% pr. million år, gældende for forskellige ordrer.
Dette særlige DNA fungerer som et værktøj til at forskning det biologiske slægtskab mellem populationer, fordi deres egenskaber gør det muligt at belyse forholdet mellem populationer der har divergeret i nyere tid, uden at skulle overveje rekombinationsfænomener og tilføje en dimension midlertidig; Til dette skal følgende huskes: to individer, hvis fælles forfader har været en kvinde, vil have mitokondrielle DNA-molekyler så forskellige som tiden er gået siden adskillelsen af forfader.
Nogle områder af DNA kan være meget lig hinanden, hvilket tillader deres klassifikation i samme klynge, som er kendt som en haplogruppe. For eksempel, Torroni et al (1993) kendetegnende de grundlæggende mitokondrielle linjer for det amerikanske kontinent, som modtog betegnelsen for haplogruppe A, B, C og D i henhold til punktmutationer af sekvensen mitokondrie. Disse mutationer skaber forskellige spaltningssteder for specifikke enzymer, som beskrevet nedenfor: i linje A er der stedet cutoff 663 for enzymet HaeIII, linje C er karakteriseret ved site 13259 for HincII, i linje D er site 5176 genkendt for AluI; denne identifikation er lavet ud fra fragment længde polymorfismer (RFLP). I tilfælde af afstamning B er der en deletion af 9 basepar ved position 8272-8289.
Den geografiske fordeling af hver slægt er blevet beskrevet som følger: slægt A er fremherskende på det amerikanske kontinent og især i Nordamerika; denne haplogruppe tilskrives dog også mesoamerikanske befolkninger. Slægter C og D optræder hovedsagelig i Sydamerika; afstamning B findes i den nordlige og sydlige region af Stillehavskysten, og Kemp et al (2010) foreslår den som en karakteristisk afstamning af familie Yuto-aztekanske og det sydvestlige U.S.A.
Derudover er der mutationer, som deles mellem individer af samme haplogruppe, hvilket har gjort det muligt at beskrive specifikke haplotyper (eller sublineages) af nogle populationer. Figur 1 viser regionerne af mitokondrielt DNA, der indeholder mutationerne for at genkende hver afstamning.
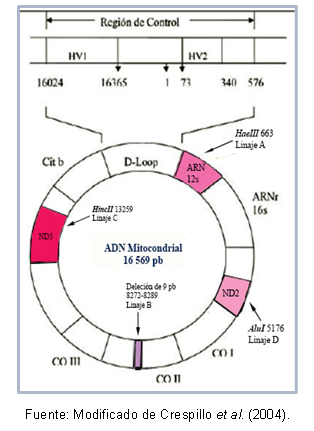
Figur 1. Mitokondrielle DNA-regioner, der kendetegner hver amerikansk stiftende slægt.
I nogle år blev kun disse fire slægter anerkendt, indtil haplogruppe X blev inkluderet for de nordlige befolkninger på kontinentet; som også er fjernt beslægtet med europæiske befolkninger. De tidligere slægter har en korrespondance med asiatiske befolkninger, selvom de observeres sjældnere end på det amerikanske kontinent; slægter A, B og C findes ikke hos moderne afrikanere og kaukasiere; og afstamning D findes også i Afrika, men forbundet med andre restriktionssteder. Med ovenstående kan vi sige, at disse haplogrupper karakteriserer de amerikanske befolkninger, og derfor er deres undersøgelse tilstrækkelig med hensyn til bosættelse og migrationer (gamle og nyere).
Måden at etablere genetiske ligheder eller forskelle mellem populationer er med genetiske afstande, som kan have en historisk forklaring, da de ændrer sig (stiger eller falder) i løbet af generationerne og kan føre os til begivenheder i historien af en befolkning for eksempel: på tidspunktet for en stor migration eller kontakt mellem to kulturer, kan vi med denne beregning skelne, hvilke mekanismer der trådte i kraft for at give os dette eller hint resultat.
Både genetisk drift og genflow har at gøre med den lethed, hvormed en gruppe, i dette tilfælde mennesker, skal bevæge sig inden for en gruppe. territorium og komme i kontakt med andre grupper. Så geografisk isolation er den genetiske afstand, der vokser med stigende geografisk afstand mellem menneskelige grupper.
For at forklare mangfoldighed i de grundlæggende mitokondrielle slægter i Amerika er der to hypotese: at dette kontinent er blevet koloniseret af flere begivenheder fra Beringia, eller at når migration først er sket, er der sket evolutionære ændringer efter koloniseringen. To indgangsveje for disse variationer til kontinentet er også forklaret, den første foreslår, at de fire grundlæggende haplogrupper uden variationer, det vil sige hver med en rodhaplotype, kunne de være ankommet lige efter det sidste istidsmaksimum eller lidt før, med datoer for 21.000 til 19.000 år siden og ville have fulgt en kystrute ved Fredelig; det andet forslag antyder, at disse intra-haplogruppe variationer allerede eksisterede i Beringia og blev bragt til den sydlige del af det amerikanske kontinent, men deres indtræden ville have været nøjagtigt i slutningen af det sidste istidsmaksimum, så ruterne inden for kontinentet allerede ville være frie, så ville disse menneskegruppers adgang være for 19 tusind siden flere år. Der er også en stor mangfoldighed af haplogruppe A og en sammensmeltningstid for den, der er kortere end resten (17 tusind år).
Den mest sandsynlige forklaring på dette er, at det skyldes sekundære udvidelser af haplogruppe A fra Beringia, længe efter afslutningen af det sidste istidsmaksimum. På trods af uoverensstemmelsen med hensyn til tidspunktet for menneskets indtog i Amerika, har genetiske undersøgelser gjort det give en vis klarhed, fordi de understøtter hypotesen om, at der var menneskegrupper på det amerikanske kontinent før Clovis; og det er fundet, at der er en adskillelse mellem forfædrene i Nordøstasien for 25-35 tusind år siden og indtoget i Amerika for 15-35 tusinde år siden. Figur 2 viser mitokondrielle haplogruppers ruter i verden og tidspunktet for divergens i år før nutiden.
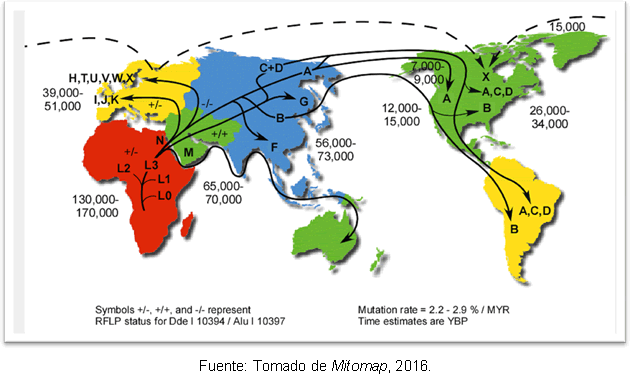
Figur 2. Kort over de forskellige spredningsruter af mitokondrielle slægter
Der er et meget nyttigt værktøj til genetisk analyse; phylogeografien. Det er de første anvendelser af molekylære undersøgelser, og med det søger vi at kunne bestemme de fylogenetiske og rumlige forhold mellem nukleotidsekvenser, i dette tilfælde DNA mitokondrie. Den rumlige fordeling kan ligne et tidsmæssigt mønster, det vil sige, at de mest geografisk fjerne DNA-sekvenser bør være de mest genetisk forskellige, såvel som DNA-sekvenser, der divergerede for længe siden, burde også være de mest forskellige genetisk. Så geografisk fjerne populationer, med ringe eller ingen genstrøm mellem dem, ville akkumulere forskelle på grund af genetisk drift og mutation, selv ved selektion; men der kan forekomme situationer, der ikke tillader drift, såsom en eller flere grundlæggereffekter eller andre mønstre for genflow.
De data, der leveres af mitokondrielle gener, har været blandt de mest nyttige i phylogeografisk forskning på grund af karakteristisk, at disse gener ikke rekombinerer, og som et resultat viser de en meget klarere fylogenetisk linje end mange nukleare gener. Den effektive populationsstørrelse beregnet med de mitokondrielle gener (og med Y-kromosomet, der fungerer på lignende måde) er omkring en fjerdedel af den, der er beregnet for de mitokondrielle gener. nukleare gener, så sker divergens næsten fire gange hurtigere end med nukleare gener, denne hurtige divergenshastighed (og genflow) kan forårsage observeret mønster af arv i disse uniparentale gener er anderledes end fylogenierne opnået med nukleare gener (som repræsenterer størstedelen af genpuljen i en individuel).
Cavalli-Sforza, L. L. (2000). Gener, folkeslag og sprog (1. udgave i Pocket Library ed.). Barcelona, Spanien: Kritisk redaktion.
Rebato, E., Susanne, C., & Chiarelli, B. (Redigeringer). (2005). At forstå biologisk antropologi: evolution og menneskelig biologi. guddommelige ord.
Torroni, A., Schurr, T. G., Cabell, M. F, Brown, M. D, Neel, J. V., Larsen, M., et al. (1993). Asiatiske affiniteter og kontinental stråling af de fire grundlæggende indianske mtDNA'er. The American Journal of Human Genetics, 53(3), 563.
(2022). mitokondrielt DNA, National Human Genome Research Institute.
-Mitokondriel DNA skematisk reference-
Crespillo, M., Paredes, M., Arimany, J., Guerrero, L., & Valverde, J. (2004). Spansk borgerkrig (1936-1939): identifikation af menneskelige rester fra massegrave i Catalonien ved mitokondriel DNA-analyse. Om en sag. Retsmedicinske notesbøger (38), 37-46.

