04/07/2021
0
Виевс

Митохондријски геном је мали, затворени кружни молекул са двоструком спиралом од 16.569 парова база (бп) који се наслеђује само од мајки на синове и ћерке (ретко). случајеви хетероплазме), тако да нема генетске рекомбинације у тренутку зачећа, она такође еволуира искључиво акумулацијом мутација у временске прилике.





Висока стопа мутације митохондријалног генома (10 до 20 пута већа од ДНК нуклеарне, у смислу гена са упоредивим функцијама) је корисно за разликовање између популација које током времена су биолошки сродне, међутим, њихова брзина мутација може бити толико брза да може доћи до појаве мутације. повратна мутација. Оцена еволуција Средња вредност овог молекула је израчуната из врста код којих су времена дивергенције од фосилних остатака била доступна, и из биодемографских или протеинских података; резултат даје 1-2% на милион година, важи за различите наруџбе.
Ова посебна ДНК функционише као алат за
истраживања биолошко сродство између популација јер њихова својства омогућавају да се разјасне односи између популација који су се у последње време разишли, без потребе да се разматрају феномени рекомбинације и додавања димензије привремени; За ово се мора имати на уму следеће: две особе чији је заједнички предак била жена ће имати молекули митохондријске ДНК различити колико је времена протекло од раздвајања предак.Неки региони ДНК могу бити веома слични једни другима, што дозвољава њихово класификација у истом кластеру, који је познат као хаплогрупа. На пример, Торрони и сарадници (1993) описали су оснивачке митохондријалне лозе за амерички континент, која је добила назив хаплогрупа А, Б, Ц и Д, према тачкастим мутацијама секвенце митохондријски. Ове мутације стварају различита места цепања за специфичне ензиме, као што је описано у наставку: у лози А постоји место цутофф 663 за ензим ХаеИИИ, лоза Ц карактерише место 13259 за ХинцИИ, у линији Д место 5176 је препознато за АлуИ; ова идентификација се врши на основу полиморфизама дужине фрагмената (РФЛП). У случају лозе Б, постоји брисање 9 парова база на позицији 8272-8289.
Географска дистрибуција сваке лозе је описана на следећи начин: лоза А преовлађује на америчком континенту и посебно у Северној Америци; међутим, ова хаплогрупа се такође приписује мезоамеричким популацијама. Линије Ц и Д појављују се углавном у Јужној Америци; лоза Б налази се у северном и јужном региону пацифичке обале, а Кемп и сарадници (2010) га предлажу као карактеристичну лозу породица Јуто-Азтекански и југозападни део САД
Поред тога, постоје мутације које се деле између појединаца исте хаплогрупе, што је омогућило да се опишу специфични хаплотипови (или подлозе) неких популација. Слика 1 приказује регионе митохондријалне ДНК који садрже мутације за препознавање сваке лозе.
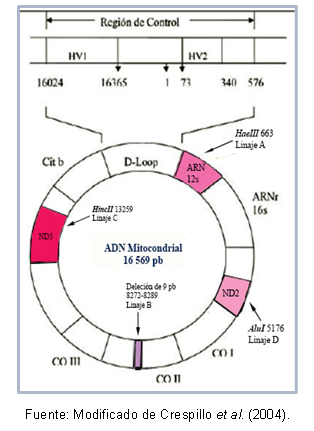
Слика 1. Региони митохондријске ДНК који су типични за сваку америчку лозу оснивача.
Неколико година само ове четири лозе су биле признате све док хаплогрупа Кс није била укључена за северне популације континента; што је такође у далекој вези са европским популацијама. Претходне лозе имају кореспонденцију са азијским популацијама, иако се примећују ређе него на америчком континенту; лозе А, Б и Ц се не налазе код модерних Африканаца и белаца; и лоза Д такође постоји у Африци, али је повезана са другим рестриктивним местима. Уз наведено, могли бисмо рећи да ове хаплогрупе карактеришу америчку популацију и стога је њихово проучавање адекватно у смислу насељавања и миграција (старих и новијих).
Начин утврђивања генетских сличности или разлика између популација је генетским удаљеностима, које могу имати а историјско објашњење, будући да се мењају (повећавају или смањују) током генерација, и могу нас довести до догађаја у историји од једног Популација на пример: у време велике сеобе или додира две културе, овим прорачуном можемо разазнати који су механизми деловали да би нам дали овај или онај резултат.
И генетски дрифт и ток гена имају везе са лакоћом са којом се група, у овом случају људска бића, мора кретати унутар групе. територија и да дође у контакт са другим групама. Дакле, географска изолација је генетска удаљеност која расте са повећањем географске удаљености између људских група.
Да објасним разноликости у оснивачким митохондријским лозама у Америци, постоје две хипотеза: да је овај континент колонизован вишеструким догађајима из Берингије, или да су се, након што је дошло до миграције, догодиле еволуционе промене након колонизације. Објашњена су и два пута уласка ових варијација на континент, први предлаже да четири оснивачке хаплогрупе без варијација, односно свака са корен хаплотипа, могли су да стигну непосредно после последњег глацијалног максимума или мало пре, са датумима од пре 21 хиљаду до 19 хиљада година и пратили би приобални пут поред Пеацефул; други предлог сугерише да су ове варијације унутар хаплогрупе већ постојале у Берингији и да су донете на југ америчког континента, али би њихов улазак био тачно на крају последњег глацијалног максимума да би руте унутар континента већ биле слободне, тада би улазак ових људских група био пре 19 хиљада године. Такође постоји велика разноликост хаплогрупе А и време спајања за њу које је краће од осталих (17 хиљада година).
Највероватније објашњење за ово је да је то последица секундарних експанзија хаплогрупе А из Берингије, дуго након завршетка последњег глацијалног максимума. Упркос нескладу у погледу времена уласка човека у Америку, генетске студије су дају извесну јасноћу, јер подржавају хипотезу да су на америчком континенту и раније постојале људске групе Цловис; и налази се да постоји раздвајање између предака североисточне Азије пре 25-35 хиљада година и уласка у Америку пре 15-35 хиљада година. На слици 2 приказани су путеви митохондријалних хаплогрупа у свету и време дивергенције у годинама пре садашњости.
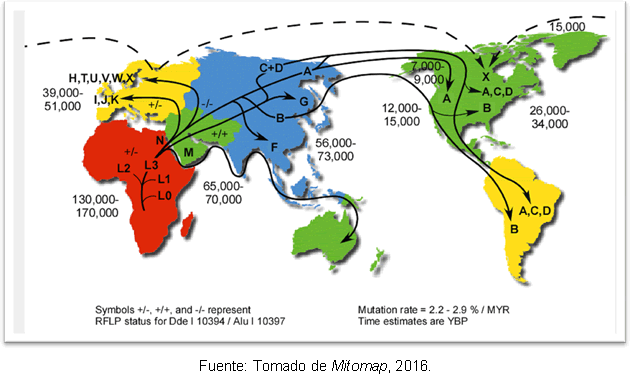
Слика 2. Мапа различитих путева дисперзије митохондријалних линија
Постоји веома користан алат за генетску анализу; филогеографија. Ово су прве примене молекуларних студија и са њима настојимо да будемо у стању да одредимо филогенетски и просторни односи између нуклеотидних секвенци, у овом случају ДНК митохондријски. Просторна дистрибуција може личити на временски образац, то јест, географски најудаљеније секвенце ДНК треба да буду најудаљеније. генетски различите, као и ДНК секвенце које су се одавно разишле, такође би требало да буду најразличитије генетски. Дакле, географски удаљене популације, са мало или нимало протока гена између њих, акумулирале би разлике услед генетског померања и мутације, чак и селекцијом; али се могу десити ситуације које не дозвољавају померање, као што су један или више ефеката оснивача или други обрасци тока гена.
Подаци добијени од митохондријалних гена били су међу најкориснијим у филогеографским истраживањима због карактеристика да се ови гени не рекомбинују и као резултат тога показују много јаснију филогенетичку линију од многих нуклеарних гена. Ефективна величина популације израчуната са митохондријалним генима (и са И хромозомом који функционише на сличан начин) је око четвртина оне израчунате за митохондријалне гене. нуклеарних гена, тада се дивергенција дешава скоро четири пута брже него код нуклеарних гена, ова брза стопа дивергенције (и проток гена) може изазвати Уочени образац наслеђивања у овим унипаренталним генима је другачији од филогенија добијених са нуклеарним генима (који представљају већину генског фонда у појединац).
Кавали-Сфорца, Л. Л. (2000). Гени, народи и језици (1. издање у Поцкет Либрари ур.). Барселона, Шпанија: Критички уводник.
Ребато, Е., Сусанне, Ц., & Цхиарелли, Б. (Уређивање.). (2005). Да разумеју биолошку антропологију: еволуција и људска биологија. Божанска реч.
Торрони, А., Сцхурр, Т. Г., Кабел, М. Ф, Браун, М. Д, Неел, Ј. В., Ларсен, М., ет ал. (1993). Азијски афинитети и континентално зрачење четири оснивача мтДНК Индијанаца. Тхе Америцан Јоурнал оф Хуман Генетицс, 53(3), 563.
(2022). митохондријалну ДНК, Национални институт за истраживање људског генома.
-Референца шеме митохондријалне ДНК-
Цреспило, М., Паредес, М., Аримани, Ј., Гуерреро, Л., & Валверде, Ј. (2004). Шпански грађански рат (1936-1939): идентификација људских остатака из масовних гробница у Каталонији анализом митохондријалне ДНК. О случају. Свеске судске медицине (38), 37-46.