
04/07/2021

Det mitokondriella genomet är en liten, sluten, dubbelspiral cirkulär molekyl på 16 569 baspar (bp) som ärvs endast från mödrar till söner och döttrar (sällan). fall av heteroplasmi), så det finns ingen genetisk rekombination vid tidpunkten för befruktningen, den utvecklas också uteslutande genom ackumulering av mutationer i väder.





Den höga mutationshastigheten i mitokondriella genomet (10 till 20 gånger den av DNA nukleära, när det gäller gener med jämförbara funktioner) är användbar för att skilja mellan populationer som över tid har varit biologiskt relaterade, men deras mutationshastighet kan vara så snabb att mutationsfenomen kan uppstå. tillbakamutation. Hastigheten på Evolution Medelvärdet för denna molekyl har beräknats från arter där avvikelsetider från fossila lämningar fanns tillgängliga och från biodemografiska eller proteindata; resultatet ger 1-2% per miljon år, giltigt för olika order.
Just detta DNA fungerar som ett verktyg för att
forskning det biologiska släktskapet mellan populationer eftersom deras egenskaper gör det möjligt att belysa sambanden mellan populationer som har divergerat på senare tid, utan att behöva ta hänsyn till rekombinationsfenomen och lägga till en dimension temporär; För detta måste följande komma ihåg: två individer vars gemensamma förfader har varit en kvinna kommer att ha mitokondriella DNA-molekyler så olika som tiden har förflutit sedan separationen av förfader.Vissa regioner av DNA kan vara mycket lika varandra, vilket tillåter deras klassificering i samma kluster, som är känd som en haplogrupp. Till exempel, Torroni et al (1993) typiserade de grundande mitokondriella linjerna för den amerikanska kontinenten, som fick valören av haplogrupper A, B, C och D, enligt punktmutationer av sekvensen mitokondriella. Dessa mutationer skapar olika klyvningsställen för specifika enzymer, som beskrivs nedan: i linje A finns stället cutoff 663 för enzymet HaeIII, härstamning C karakteriseras av plats 13259 för HincII, i linje D är plats 5176 igenkänd för AluI; denna identifiering görs från fragmentlängdspolymorfismer (RFLP). I fallet med härstamning B finns det en radering av 9 baspar vid position 8272-8289.
Den geografiska fördelningen av varje släktlinje har beskrivits enligt följande: släktlinje A är dominerande på den amerikanska kontinenten och särskilt i Nordamerika; denna haplogrupp tillskrivs emellertid också mesoamerikanska befolkningar. Linjerna C och D förekommer huvudsakligen i Sydamerika; härstamning B finns i den norra och södra delen av Stillahavskusten, och Kemp et al (2010) föreslår den som en karakteristisk härstamning för familj Yuto-aztekanska och sydvästra U.S.A.
Dessutom finns det mutationer som delas mellan individer av samma haplogrupp, vilket har gjort det möjligt att beskriva specifika haplotyper (eller sublineages) av vissa populationer. Figur 1 visar de regioner av mitokondriella DNA: t som innehåller mutationerna för att känna igen varje härstamning.
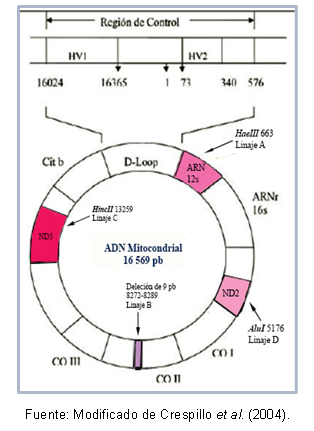
Figur 1. Mitokondriella DNA-regioner som kännetecknar varje amerikansk grundlinje.
Under några år erkändes endast dessa fyra linjer tills haplogrupp X inkluderades för de nordliga befolkningarna på kontinenten; som också är avlägset släkt med europeiska befolkningar. De tidigare linjerna har en överensstämmelse med asiatiska befolkningar, även om de observeras mer sällan än på den amerikanska kontinenten; linjerna A, B och C finns inte hos moderna afrikaner och kaukasier; och linje D finns också i Afrika, men associerad med andra restriktionsställen. Med ovanstående kan vi säga att dessa haplogrupper kännetecknar de amerikanska populationerna och därför är deras studie adekvat när det gäller bosättning och migrationer (gamla och nyare).
Sättet att fastställa genetiska likheter eller skillnader mellan populationer är med genetiska avstånd, som kan ha en historisk förklaring, eftersom de förändras (ökar eller minskar) under generationerna och kan leda oss till händelser i historien av en befolkning till exempel: vid tidpunkten för en stor migration eller kontakt mellan två kulturer, med denna beräkning kan vi urskilja vilka mekanismer som trädde i kraft för att ge oss det eller det resultatet.
Både genetisk drift och genflöde har att göra med hur lätt en grupp, i detta fall människor, måste röra sig inom en grupp. territorium och att komma i kontakt med andra grupper. Så, geografisk isolering är det genetiska avståndet som växer med ökande geografiska avstånd mellan mänskliga grupper.
För att förklara mångfald i de grundande mitokondriella linjerna i Amerika finns det två hypotes: att denna kontinent har koloniserats av flera händelser från Beringia, eller att när migration har inträffat har evolutionära förändringar inträffat efter koloniseringen. Två inträdesvägar för dessa variationer till kontinenten förklaras också, den första föreslår att de fyra grundande haplogrupperna utan variationer, det vill säga var och en med en rothaplotyp, kunde de ha anlänt strax efter det sista istidsmaximum eller lite tidigare, med datum för 21 tusen till 19 tusen år sedan och skulle ha följt en kustväg av Fredlig; det andra förslaget antyder att dessa intrahaplogruppsvariationer redan existerade i Beringia och fördes till södra den amerikanska kontinenten men deras inträde skulle ha varit exakt i slutet av Last Glacial Maximum så att rutterna inom kontinenten redan skulle vara fria, då skulle inträdet för dessa mänskliga grupper vara 19 tusen sedan år. Det finns också en stor mångfald av haplogrupp A och en koalescenstid för den som är kortare än resten (17 tusen år).
Den mest troliga förklaringen till detta är att det beror på sekundära expansioner av haplogrupp A från Beringia, långt efter slutet av det sista istidsmaximum. Trots diskrepansen när det gäller tidpunkten för människans inträde i Amerika, har genetiska studier gjort det ge lite klarhet, eftersom de stödjer hypotesen att det fanns mänskliga grupper på den amerikanska kontinenten tidigare Clovis; och man har funnit att det finns en separation mellan förfäderna i Nordostasien för 25-35 tusen år sedan och inträdet i Amerika för 15-35 tusen år sedan. Figur 2 visar vägarna för de mitokondriella haplogrupperna i världen och tidpunkten för divergensen år före nutid.
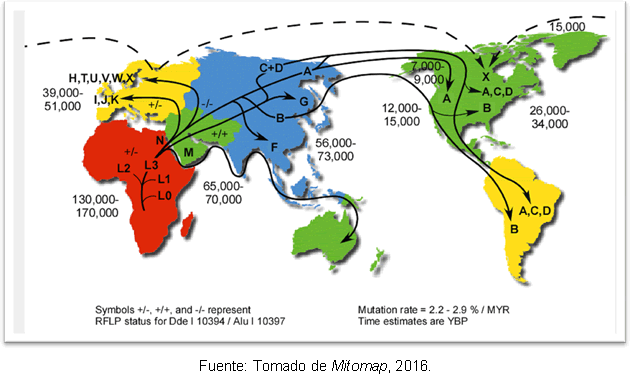
Figur 2. Karta över de olika spridningsvägarna för mitokondriella linjer
Det finns ett mycket användbart verktyg för genetisk analys; fylogeografin. Dessa är de första tillämpningarna av molekylära studier, och med det försöker vi kunna avgöra de fylogenetiska och rumsliga förhållandena mellan nukleotidsekvenser, i detta fall DNA mitokondriella. Den rumsliga fördelningen kan likna ett tidsmönster, det vill säga de mest geografiskt avlägsna DNA-sekvenserna bör vara de mest genetiskt olika, liksom DNA-sekvenser som avvikit för länge sedan borde också vara de mest olika genetiskt. Så geografiskt avlägsna populationer, med lite eller inget genflöde mellan dem, skulle ackumulera skillnader på grund av genetisk drift och mutation, även genom selektion; men situationer kan uppstå som inte tillåter drift, såsom en eller flera grundareffekter eller andra genflödesmönster.
Data som tillhandahålls av mitokondriella gener har varit bland de mest användbara inom fylogeografiforskning på grund av egenskaper som dessa gener har att inte rekombinera och som ett resultat visar de en mycket tydligare fylogenetisk linje än många kärngener. Den effektiva populationsstorleken beräknad med mitokondriella gener (och med Y-kromosomen som fungerar på liknande sätt) är ungefär en fjärdedel av den som beräknats för mitokondriella gener. kärngener, då divergens sker nästan fyra gånger snabbare än med kärngener, kan denna snabba divergenshastighet (och genflöde) orsaka observerade arvsmönster i dessa uniparentala gener skiljer sig från fylogenierna som erhålls med nukleära gener (som representerar majoriteten av genpoolen i en enskild).
Cavalli-Sforza, L. L. (2000). Gener, folk och språk (1st Edition i Pocket Library ed.). Barcelona, Spanien: Kritisk redaktion.
Rebato, E., Susanne, C., & Chiarelli, B. (Redigeringar.). (2005). Att förstå biologisk antropologi: evolution och mänsklig biologi. Gudomligt Ord.
Torroni, A., Schurr, T. G., Cabell, M. F, Brown, M. D, Neel, J. V., Larsen, M., et al. (1993). Asiatisk affinitet och kontinental strålning av de fyra grundande indianska mtDNA: erna. American Journal of Human Genetics, 53(3), 563.
(2022). mitokondriellt DNA, National Human Genome Research Institute.
-Mitokondriell DNA schematisk referens-
Crespillo, M., Paredes, M., Arimany, J., Guerrero, L., & Valverde, J. (2004). Spanska inbördeskriget (1936-1939): identifiering av mänskliga kvarlevor från massgravar i Katalonien genom mitokondriell DNA-analys. Om ett fall. Rättsmedicinska anteckningsböcker (38), 37-46.

