
13/11/2021

จีโนมของไมโตคอนเดรียเป็นโมเลกุลทรงกลมขนาดเล็กแบบปิดและเป็นเกลียวคู่ที่มีจำนวนคู่เบส 16,569 คู่เบส (bp) ซึ่งสืบทอดมาจากแม่ถึงลูกชายและลูกสาวเท่านั้น (หายาก) กรณีของ heteroplasmy) ดังนั้นจึงไม่มีการรวมตัวของยีนในช่วงเวลาของการปฏิสนธิ มันยังวิวัฒนาการโดยการสะสมของการกลายพันธุ์ใน สภาพอากาศ.





อัตราการกลายพันธุ์ของยีน mitochondrial สูง (10 ถึง 20 เท่าของ ดีเอ็นเอ นิวเคลียร์ในแง่ของยีนที่มีหน้าที่เทียบเคียง) มีประโยชน์สำหรับการแยกความแตกต่างระหว่างประชากรที่เมื่อเวลาผ่านไป มีความเกี่ยวข้องทางชีววิทยา อย่างไรก็ตาม อัตราการกลายพันธุ์ของพวกมันเร็วมากจนปรากฏการณ์การกลายพันธุ์สามารถเกิดขึ้นได้ ย้อนกลับ อัตราของ วิวัฒนาการ ค่าเฉลี่ยของโมเลกุลนี้คำนวณจากสปีชีส์ที่มีเวลาความแตกต่างจากซากฟอสซิล และจากข้อมูลทางประชากรศาสตร์หรือโปรตีน ผลลัพธ์ให้ 1-2% ต่อล้านปี ใช้ได้สำหรับคำสั่งซื้อที่แตกต่างกัน
ดีเอ็นเอเฉพาะนี้ทำหน้าที่เป็นเครื่องมือในการ การวิจัย เครือญาติทางชีววิทยาระหว่างประชากรเนื่องจากคุณสมบัติของพวกมันช่วยให้เข้าใจถึงความสัมพันธ์ระหว่างประชากร ที่ได้เบี่ยงออกไปเมื่อไม่นานนี้ โดยไม่ต้องคำนึงถึงปรากฏการณ์การรวมตัวกันใหม่และเพิ่มมิติ ชั่วคราว; ทั้งนี้ พึงระลึกไว้เสมอว่า บุคคล ๒ คน ที่บรรพบุรุษร่วมกันเป็นสตรีย่อมมี โมเลกุล DNA ของไมโตคอนเดรียแตกต่างไปตามกาลเวลานับตั้งแต่การแยกตัวของ บรรพบุรุษ.
DNA บางภูมิภาคอาจมีความคล้ายคลึงกันมาก ทำให้ การจำแนกประเภท ในคลัสเตอร์เดียวกัน ซึ่งเรียกว่าแฮปโลกรุ๊ป ตัวอย่างเช่น Torroni et al (1993) ได้ระบุถึงการสืบเชื้อสายของไมโตคอนเดรียที่ก่อตัวในทวีปอเมริกา ซึ่งได้รับการตั้งชื่อของกลุ่มแฮปโลกรุ๊ป A, B, C และ D ตามการกลายพันธุ์แบบจุดของลำดับ ไมโตคอนเดรีย การกลายพันธุ์เหล่านี้สร้างตำแหน่งการแตกแยกที่แตกต่างกันสำหรับเอ็นไซม์เฉพาะ ดังที่อธิบายไว้ด้านล่าง: ในเชื้อสาย A มีไซต์ คัทออฟ 663 สำหรับเอ็นไซม์ HaeIII เชื้อสาย C มีลักษณะเฉพาะโดยไซต์ 13259 สำหรับ HincII ในเชื้อสาย D ไซต์ 5176 เป็นที่รู้จักสำหรับ AluI; การระบุนี้ทำมาจากความแตกต่างของความยาวชิ้นส่วน (RFLP) ในกรณีของเชื้อสาย B มีการลบคู่ฐาน 9 คู่ที่ตำแหน่ง 8272-8289
มีการอธิบายการกระจายทางภูมิศาสตร์ของแต่ละเชื้อสายดังนี้: เชื้อสาย A มีความสำคัญในทวีปอเมริกาและโดยเฉพาะอย่างยิ่งในอเมริกาเหนือ อย่างไรก็ตาม haplogroup นี้มาจากประชากร Mesoamerican ด้วย Lineages C และ D ส่วนใหญ่ปรากฏในอเมริกาใต้ เชื้อสาย B พบได้ในพื้นที่ภาคเหนือและภาคใต้ของชายฝั่งแปซิฟิก และ Kemp et al (2010) เสนอให้เป็นเชื้อสายที่มีลักษณะเฉพาะของ ตระกูล Yuto-Aztecan และทางตะวันตกเฉียงใต้ของสหรัฐอเมริกา
นอกจากนี้ยังมีการกลายพันธุ์ที่ใช้ร่วมกันระหว่างบุคคลในกลุ่มแฮ็ปโลกรุ๊ปเดียวกัน ซึ่งทำให้สามารถอธิบายลักษณะแฮ็ปโลไทป์เฉพาะ (หรือสายย่อย) ของประชากรบางกลุ่มได้ รูปที่ 1 แสดงบริเวณของ DNA ของไมโตคอนเดรียที่มีการกลายพันธุ์เพื่อจำแนกแต่ละเชื้อสาย
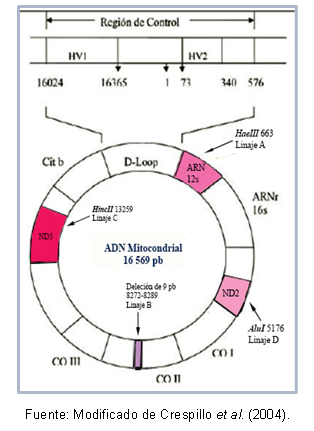
รูปที่ 1. ภูมิภาค DNA ของไมโตคอนเดรียที่แสดงถึงสายเลือดผู้ก่อตั้งอเมริกันแต่ละสาย
หลายปีที่ผ่านมามีเพียงสี่เชื้อสายเท่านั้นที่ได้รับการยอมรับจนกระทั่งรวมกลุ่ม haplogroup X ไว้สำหรับประชากรทางตอนเหนือของทวีป ซึ่งสัมพันธ์กับประชากรยุโรปอย่างห่างไกล เชื้อสายก่อนหน้านี้มีความเกี่ยวเนื่องกับประชากรเอเชีย แม้ว่าจะมีการพบเห็นน้อยกว่าในทวีปอเมริกาก็ตาม เชื้อสาย A, B และ C ไม่พบในชาวแอฟริกันและคอเคเชี่ยนสมัยใหม่ และเชื้อสาย D ยังมีอยู่ในแอฟริกา แต่เกี่ยวข้องกับไซต์ข้อจำกัดอื่นๆ จากที่กล่าวมาข้างต้น เราสามารถพูดได้ว่ากลุ่มแฮ็ปโลกรุ๊ปเหล่านี้เป็นลักษณะของประชากรอเมริกัน ดังนั้นการศึกษาของพวกมันจึงเพียงพอในแง่ของการตั้งถิ่นฐานและการอพยพ (ทั้งเก่าและใหม่)
วิธีในการสร้างความเหมือนหรือความแตกต่างทางพันธุกรรมระหว่างประชากรคือกับระยะทางทางพันธุกรรม ซึ่งสามารถมี คำอธิบายทางประวัติศาสตร์ เนื่องจากมีการเปลี่ยนแปลง (เพิ่มขึ้นหรือลดลง) หลายชั่วอายุคน และสามารถนำเราไปสู่เหตุการณ์ในประวัติศาสตร์ได้ หนึ่ง ประชากร ตัวอย่างเช่น ในช่วงเวลาของการย้ายถิ่นครั้งใหญ่หรือการติดต่อระหว่างสองวัฒนธรรม ด้วยการคำนวณนี้ เราสามารถแยกแยะได้ว่ากลไกใดเข้ามาดำเนินการเพื่อให้ผลลัพธ์นี้หรือผลลัพธ์นั้นแก่เรา
ทั้งการเคลื่อนตัวของยีนและการไหลของยีนนั้นเกี่ยวข้องกับความสะดวกที่กลุ่ม ในกรณีนี้ มนุษย์ ต้องเคลื่อนไหวภายในกลุ่ม อาณาเขต และติดต่อกับกลุ่มอื่นๆ ดังนั้น การแยกตัวทางภูมิศาสตร์คือระยะห่างทางพันธุกรรมที่เพิ่มขึ้นพร้อมกับระยะห่างทางภูมิศาสตร์ระหว่างกลุ่มมนุษย์ที่เพิ่มขึ้น
เพื่ออธิบาย ความหลากหลาย ในการก่อตั้งสายเลือดไมโตคอนเดรียในอเมริกามี 2 แบบ สมมติฐาน: ทวีปนี้ถูกล่าอาณานิคมด้วยเหตุการณ์หลายเหตุการณ์จาก Beringia หรือเมื่อเกิดการอพยพ การเปลี่ยนแปลงทางวิวัฒนาการก็เกิดขึ้นหลังจากการล่าอาณานิคม มีการอธิบายเส้นทางเข้าสู่การเปลี่ยนแปลงเหล่านี้สองเส้นทางไปยังทวีปด้วย ประการแรกเสนอว่ากลุ่มแฮปโลกรุ๊ปที่ก่อตั้งสี่กลุ่มโดยไม่มีการเปลี่ยนแปลง กล่าวคือ แต่ละกลุ่มมี รากแบบแฮปโลไทป์ พวกมันอาจมาถึงหลัง Last Glacial Maximum หรือก่อนหน้านั้นเล็กน้อย โดยมีวันที่เมื่อ 21,000 ถึง 19,000 ปีก่อน และจะไปตามเส้นทางชายฝั่งโดย สงบ; ข้อเสนอที่สองชี้ให้เห็นว่ารูปแบบภายในกลุ่มแฮปโลกรุ๊ปเหล่านี้มีอยู่แล้วในเบรินเจียและถูกนำตัวไปทางใต้ของทวีปอเมริกา แต่การเข้ามาของพวกเขาน่าจะเป็น ตรงปลายสุดของ Last Glacial Maximum ดังนั้นเส้นทางภายในทวีปจะว่างอยู่แล้ว ดังนั้นการเข้ามาของกลุ่มมนุษย์เหล่านี้จะเป็น 19,000 ที่แล้ว ปี. นอกจากนี้ยังมีกลุ่ม haplogroup A ที่หลากหลายและเวลาในการรวมตัวกันซึ่งสั้นกว่าส่วนที่เหลือ (17,000 ปี)
คำอธิบายที่น่าจะเป็นไปได้มากที่สุดสำหรับเรื่องนี้ก็คือ เนื่องมาจากการขยายลำดับรองของ haplogroup A จาก Beringia หลังจากสิ้นสุด Last Glacial Maximum เป็นเวลานาน แม้จะมีความคลาดเคลื่อนเกี่ยวกับเวลาที่มนุษย์เข้าสู่ทวีปอเมริกา แต่การศึกษาทางพันธุกรรมก็มี ให้ความกระจ่างบ้างเพราะสนับสนุนสมมติฐานว่าเคยมีกลุ่มมนุษย์ในทวีปอเมริกามาก่อน โคลวิส; และพบว่ามีการแตกแยกระหว่างบรรพบุรุษของเอเชียตะวันออกเฉียงเหนือเมื่อ 25-35,000 ปีก่อนกับการเข้าสู่อเมริกาเมื่อ 15-35,000 ปีก่อน รูปที่ 2 แสดงเส้นทางของกลุ่ม haplogroups ของไมโตคอนเดรียในโลกและเวลาของความแตกต่างในปีก่อนปัจจุบัน
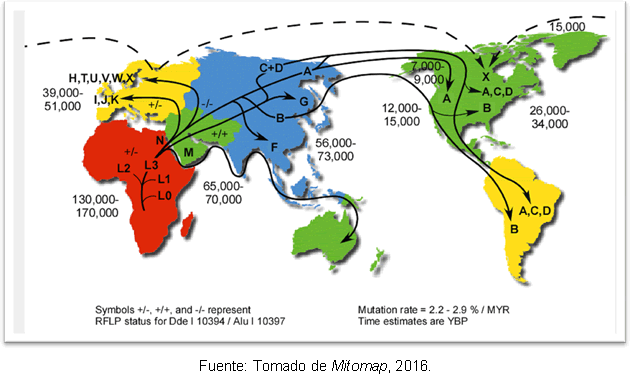
รูปที่ 2 แผนที่เส้นทางการแพร่กระจายของสายเลือดไมโตคอนเดรียต่างๆ
มีเครื่องมือที่มีประโยชน์มากสำหรับการวิเคราะห์ทางพันธุกรรม สายวิวัฒนาการ นี่เป็นแอปพลิเคชั่นแรกของการศึกษาระดับโมเลกุล และด้วยเหตุนี้ เราจึงพยายามที่จะระบุ ความสัมพันธ์ทางสายวิวัฒนาการและเชิงพื้นที่ระหว่างลำดับนิวคลีโอไทด์ ในกรณีนี้คือ DNA ไมโตคอนเดรีย การกระจายเชิงพื้นที่อาจคล้ายกับรูปแบบชั่วขณะ กล่าวคือ ลำดับดีเอ็นเอที่อยู่ไกลที่สุดทางภูมิศาสตร์ควรเป็นลำดับมากที่สุด ทางพันธุกรรมที่แตกต่างกันไป เช่นเดียวกับลำดับดีเอ็นเอที่แยกจากกันเมื่อนานมาแล้วก็ควรจะแตกต่างกันมากที่สุด ทางพันธุกรรม ดังนั้น ประชากรที่อยู่ห่างไกลตามภูมิศาสตร์ ซึ่งมีการไหลของยีนเพียงเล็กน้อยหรือไม่มีเลยระหว่างกัน จะสะสมความแตกต่างเนื่องจากการเคลื่อนตัวของยีนและการกลายพันธุ์ แม้กระทั่งโดยการคัดเลือก แต่สถานการณ์อาจเกิดขึ้นที่ไม่อนุญาตให้ล่องลอยได้ เช่น ผลกระทบของผู้ก่อตั้งอย่างน้อยหนึ่งรูปแบบหรือรูปแบบอื่น ๆ ของการไหลของยีน
ข้อมูลที่ได้จากยีนไมโทคอนเดรียเป็นประโยชน์มากที่สุดในการวิจัยสายวิวัฒนาการเนื่องจาก ลักษณะเฉพาะที่ยีนเหล่านี้ไม่มีการรวมตัวใหม่ และเป็นผลให้ยีนเหล่านี้แสดงเส้นสายวิวัฒนาการที่ชัดเจนกว่าหลายๆ ยีนนิวเคลียร์ ขนาดประชากรที่มีประสิทธิภาพคำนวณด้วยยีนยล (และด้วยการทำงานของโครโมโซม Y ในลักษณะเดียวกัน) ประมาณหนึ่งในสี่ของจำนวนที่คำนวณสำหรับยีนยล ยีนนิวเคลียร์ จากนั้นไดเวอร์เจนซ์จะเกิดขึ้นเร็วกว่ายีนนิวเคลียร์เกือบสี่เท่า อัตราการเปลี่ยนแปลงอย่างรวดเร็วนี้ (และการไหลของยีน) อาจทำให้เกิด รูปแบบการสืบทอดที่สังเกตได้ในยีนที่มีพ่อแม่เดียวเหล่านี้แตกต่างจากสายวิวัฒนาการที่ได้รับจากยีนนิวเคลียร์ (ซึ่งเป็นตัวแทนของกลุ่มยีนส่วนใหญ่ใน รายบุคคล).
คาวาลลี-สฟอร์ซา, แอล. ล. (2000). ยีน ประชาชน และภาษา (ฉบับที่ 1 ใน Pocket Library ed.) บาร์เซโลนา สเปน: บทบรรณาธิการที่สำคัญ.
Rebato, E., Susanne, C. และ Chiarelli, B. (แก้ไข) (2005). เพื่อทำความเข้าใจมานุษยวิทยาชีวภาพ: วิวัฒนาการและชีววิทยาของมนุษย์ คำศักดิ์สิทธิ์.
Torroni, A., Schurr, ที. จี, คาเบลล์, เอ็ม. เอฟ บราวน์ เอ็ม ดี, นีล, เจ. V., Larsen, M., et al. (1993). ความสัมพันธ์แบบเอเชียและการแผ่รังสีของทวีปของ mtDNAs ของชนพื้นเมืองอเมริกันทั้งสี่กลุ่ม. American Journal of Human Genetics, 53(3), 563.
(2022). ดีเอ็นเอของไมโตคอนเดรีย,สถาบันวิจัยจีโนมมนุษย์แห่งชาติ.
-การอ้างอิงแผนผังดีเอ็นเอของไมโตคอนเดรีย-
Crespillo, M., Paredes, M., Arimany, J., Guerrero, L., & Valverde, J. (2004). สงครามกลางเมืองสเปน (พ.ศ. 2479-2482): การระบุซากศพมนุษย์จากหลุมศพขนาดใหญ่ในคาตาโลเนียโดยการวิเคราะห์ดีเอ็นเอของไมโตคอนเดรีย เกี่ยวกับคดี. สมุดบันทึกนิติเวช (38), 37-46.