04/07/2021
0
Დათვალიერება

მიტოქონდრიული გენომი არის პატარა, დახურული, ორმაგი სპირალის წრიული მოლეკულა 16,569 ბაზის წყვილით (bp), რომელიც მემკვიდრეობით გადადის მხოლოდ დედებიდან ვაჟებსა და ქალიშვილებზე (იშვიათად). ჰეტეროპლაზმის შემთხვევები), ასე რომ, ჩასახვის დროს არ არსებობს გენეტიკური რეკომბინაცია, ის ასევე ვითარდება ექსკლუზიურად მუტაციების დაგროვებით ამინდი.





მიტოქონდრიული გენომის მუტაციის მაღალი მაჩვენებელი (10-დან 20-ჯერ მეტი დნმ ბირთვული, შესადარებელი ფუნქციების მქონე გენების თვალსაზრისით) სასარგებლოა პოპულაციების დიფერენცირებისთვის, რომლებიც დროთა განმავლობაში ისინი ბიოლოგიურად იყვნენ დაკავშირებული, თუმცა მათი მუტაციის სიჩქარე შეიძლება იყოს იმდენად სწრაფი, რომ მუტაციის ფენომენები შეიძლება მოხდეს. უკუმუტაცია. მაჩვენებელი ევოლუცია ამ მოლეკულის საშუალო მაჩვენებელი გამოითვლება იმ სახეობებიდან, რომლებშიც შესაძლებელი იყო ნამარხი ნაშთებისგან განსხვავებების დრო და ბიოდემოგრაფიული ან ცილების მონაცემებიდან; შედეგი იძლევა 1-2% მილიონ წელიწადში, მოქმედებს სხვადასხვა შეკვეთებისთვის.
ეს კონკრეტული დნმ ფუნქციონირებს როგორც ინსტრუმენტი კვლევა ბიოლოგიური ნათესაობა პოპულაციებს შორის, რადგან მათი თვისებები იძლევა პოპულაციებს შორის ურთიერთობების გარკვევის საშუალებას რომლებიც განსხვავდებიან ბოლო დროს, რეკომბინაციის ფენომენების განხილვისა და განზომილების დამატების გარეშე დროებითი; ამისთვის უნდა გავითვალისწინოთ შემდეგი: ეყოლება ორ ინდივიდს, რომელთა საერთო წინაპარი ქალი ყოფილა მიტოქონდრიული დნმ-ის მოლეკულები ისეთივე განსხვავებულია, როგორიც დრო გავიდა გამოყოფის შემდეგ წინაპარი.
დნმ-ის ზოგიერთი რეგიონი შეიძლება ძალიან ჰგავს ერთმანეთს, რაც მათ საშუალებას აძლევს კლასიფიკაცია იმავე კლასტერში, რომელიც ცნობილია როგორც ჰაპლოჯგუფი. მაგალითად, ტორონი და სხვები (1993) ასახავდნენ ამერიკის კონტინენტის დამფუძნებელ მიტოქონდრიულ შტოებს. რომელმაც მიიღო ჰაპლოჯგუფების A, B, C და D ნომინალი მიმდევრობის წერტილოვანი მუტაციების მიხედვით მიტოქონდრიული. ეს მუტაციები ქმნის განსხვავებულ დაშლის ადგილებს კონკრეტული ფერმენტებისთვის, როგორც ეს აღწერილია ქვემოთ: A ხაზით არის ადგილი cutoff 663 ფერმენტისთვის HaeIII, C ხაზი ხასიათდება 13259 ადგილით HincII-სთვის, შტოში D ადგილი 5176 აღიარებულია AluI; ეს იდენტიფიკაცია კეთდება ფრაგმენტის სიგრძის პოლიმორფიზმისგან (RFLP). B ხაზის შემთხვევაში, ხდება 9 ბაზის წყვილის წაშლა 8272-8289 პოზიციაზე.
თითოეული ხაზის გეოგრაფიული გავრცელება აღწერილია შემდეგნაირად: A ხაზი გაბატონებულია ამერიკის კონტინენტზე და განსაკუთრებით ჩრდილოეთ ამერიკაში; თუმცა, ეს ჰაპლოჯგუფი ასევე მიეკუთვნება მესოამერიკელ პოპულაციებს. შტოები C და D ძირითადად სამხრეთ ამერიკაში ჩნდება; B ხაზი გვხვდება წყნარი ოკეანის სანაპიროს ჩრდილოეთ და სამხრეთ რეგიონში და Kemp et al (2010) გვთავაზობს მას, როგორც დამახასიათებელ შტოს. ოჯახი იუტო-აცტეკანი და სამხრეთ-დასავლეთი აშშ.
გარდა ამისა, არსებობს მუტაციები, რომლებიც საერთოა ერთი და იგივე ჰაპლოჯგუფის ინდივიდებს შორის, რამაც შესაძლებელი გახადა ზოგიერთი პოპულაციის კონკრეტული ჰაპლოტიპების (ან ქვეხაზების) აღწერა. სურათი 1 გვიჩვენებს მიტოქონდრიული დნმ-ის რეგიონებს, რომლებიც შეიცავს მუტაციებს თითოეული ხაზის ამოცნობისთვის.
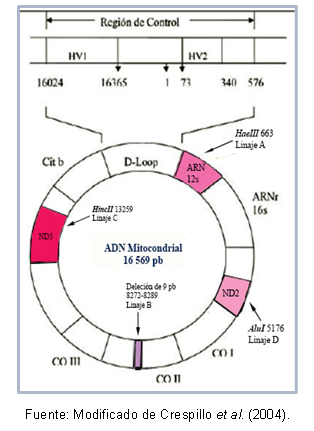
Ფიგურა 1. მიტოქონდრიული დნმ-ის რეგიონები, რომლებიც ახასიათებს თითოეულ ამერიკულ დამფუძნებელ შტოს.
რამდენიმე წლის განმავლობაში მხოლოდ ეს ოთხი ხაზი იყო აღიარებული, სანამ ჰაპლოჯგუფი X არ შედიოდა კონტინენტის ჩრდილოეთ პოპულაციებში; რომელიც ასევე შორს არის დაკავშირებული ევროპულ პოპულაციებთან. წინა გვარებს აქვთ მიმოწერა აზიურ პოპულაციებთან, თუმცა ისინი ნაკლებად ხშირად შეინიშნება, ვიდრე ამერიკის კონტინენტზე; შტოები A, B და C არ გვხვდება თანამედროვე აფრიკელებსა და კავკასიელებში; და ხაზი D ასევე არსებობს აფრიკაში, მაგრამ დაკავშირებულია სხვა შეზღუდვის ადგილებთან. ზემოაღნიშნულით შეიძლება ითქვას, რომ ეს ჰაპლოჯგუფები ახასიათებს ამერიკულ პოპულაციებს და შესაბამისად მათი შესწავლა ადეკვატურია განსახლებისა და მიგრაციების თვალსაზრისით (ძველი და უახლესი).
პოპულაციებს შორის გენეტიკური მსგავსების ან განსხვავებების დადგენის გზა არის გენეტიკური მანძილი, რომელსაც შეიძლება ჰქონდეს ა ისტორიული ახსნა, რადგან ისინი იცვლება (იზრდება ან მცირდება) თაობების განმავლობაში და შეიძლება მიგვიყვანოს ისტორიის მოვლენებამდე ერთის მოსახლეობა მაგალითად: ორ კულტურას შორის დიდი მიგრაციის ან კონტაქტის დროს, ამ გაანგარიშებით ჩვენ შეგვიძლია გავიგოთ, რა მექანიზმები ამოქმედდა, რომ მოგვცეს ესა თუ ის შედეგი.
გენეტიკური დრიფტიც და გენის ნაკადიც დაკავშირებულია იმ სიმარტივასთან, რომლითაც ჯგუფს, ამ შემთხვევაში ადამიანებს, უწევს ჯგუფში გადაადგილება. ტერიტორია და დაუკავშირდეს სხვა ჯგუფებს. ასე რომ, გეოგრაფიული იზოლაცია არის გენეტიკური მანძილი, რომელიც იზრდება ადამიანთა ჯგუფებს შორის გეოგრაფიული მანძილის მატებასთან ერთად.
ასახსნელად მრავალფეროვნება ამერიკაში დამფუძნებელ მიტოქონდრიულ ხაზებში ორია ჰიპოთეზა: რომ ეს კონტინენტი იყო კოლონიზებული ბერინგიის მრავალი მოვლენის შედეგად, ან რომ მას შემდეგ რაც მოხდა მიგრაცია, კოლონიზაციის შემდეგ მოხდა ევოლუციური ცვლილებები. ასევე ახსნილია ამ ვარიაციების კონტინენტზე შესვლის ორი მარშრუტი, პირველი ვარაუდობს, რომ ოთხი დამფუძნებელი ჰაპლოჯგუფი ვარიაციების გარეშე, ანუ თითოეულს აქვს ფესვის ჰაპლოტიპი, ისინი შეიძლება ჩამოსულიყვნენ ბოლო მყინვარული მაქსიმუმის შემდეგ ან ცოტა ადრე, თარიღებით 21 ათასიდან 19 ათასი წლის წინ და გაჰყვებოდნენ სანაპირო მარშრუტს მშვიდობიანი; მეორე წინადადება ვარაუდობს, რომ ეს შიდაჰაპლოჯგუფური ვარიაციები უკვე არსებობდა ბერინგიაში და მიტანილი იყო ამერიკის კონტინენტის სამხრეთით, მაგრამ მათი შესვლა იქნებოდა ზუსტად ბოლო მყინვარული მაქსიმუმის ბოლოს, რათა კონტინენტის შიგნით მარშრუტები უკვე თავისუფალი ყოფილიყო, მაშინ ამ ადამიანთა ჯგუფების შესვლა იქნებოდა 19 ათასი წინ. წლები. ასევე არსებობს ჰაპლოჯგუფ A-ს დიდი მრავალფეროვნება და მისი შერწყმის დრო, რომელიც დანარჩენზე უფრო მოკლეა (17 ათასი წელი).
ამის ყველაზე სავარაუდო ახსნა არის ის, რომ ეს გამოწვეულია A ჰაპლოჯგუფის მეორადი გაფართოებით ბერინგიიდან, ბოლო მყინვარული მაქსიმუმის დასრულებიდან დიდი ხნის შემდეგ. მიუხედავად შეუსაბამობისა ადამიანის ამერიკაში შესვლის დროში, გენეტიკური კვლევები გამოვლინდა მისცეს გარკვეული სიცხადე, რადგან ისინი მხარს უჭერენ ჰიპოთეზას, რომ ადრე არსებობდა ადამიანთა ჯგუფები ამერიკის კონტინენტზე კლოვისი; და აღმოჩნდა, რომ 25-35 ათასი წლის წინ ჩრდილო-აღმოსავლეთ აზიის წინაპრებსა და ამერიკაში 15-35 ათასი წლის წინ შემოსვლას შორის არის გამიჯვნა. სურათი 2 გვიჩვენებს მიტოქონდრიული ჰაპლოჯგუფების მარშრუტებს მსოფლიოში და განსხვავების დრო აწმყომდე წლების განმავლობაში.
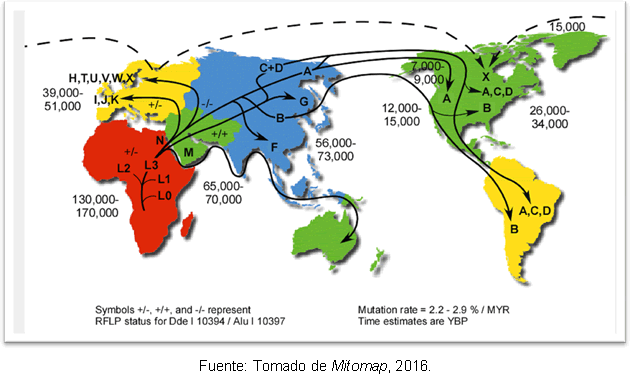
სურათი 2. მიტოქონდრიული ხაზის დისპერსიის სხვადასხვა მარშრუტების რუკა
არსებობს ძალიან სასარგებლო ინსტრუმენტი გენეტიკური ანალიზისთვის; ფილოგეოგრაფია. ეს არის მოლეკულური კვლევების პირველი აპლიკაციები და მასთან ერთად ჩვენ ვცდილობთ შევძლოთ განვსაზღვროთ ფილოგენეტიკური და სივრცითი ურთიერთობები ნუკლეოტიდურ თანმიმდევრობებს შორის, ამ შემთხვევაში დნმ-ს შორის მიტოქონდრიული. სივრცითი განაწილება შეიძლება ჰგავდეს დროებით ნიმუშს, ანუ გეოგრაფიულად ყველაზე შორეული დნმ-ის თანმიმდევრობა უნდა იყოს ყველაზე მეტად გენეტიკურად განსხვავებული, ისევე როგორც დნმ-ის თანმიმდევრობები, რომლებიც დიდი ხნის წინ განსხვავდებოდა, ასევე უნდა იყოს ყველაზე განსხვავებული გენეტიკურად. ასე რომ, გეოგრაფიულად დაშორებული პოპულაციები, მათ შორის გენის მცირე ნაკადით ან საერთოდ არ არის, აგროვებენ განსხვავებებს გენეტიკური დრიფტისა და მუტაციის გამო, თუნდაც შერჩევით; მაგრამ შეიძლება მოხდეს სიტუაციები, რომლებიც არ იძლევა დრიფტის საშუალებას, როგორიცაა ერთი ან მეტი დამფუძნებელი ეფექტი ან გენის ნაკადის სხვა ნიმუშები.
მიტოქონდრიული გენების მიერ მოწოდებული მონაცემები ერთ-ერთი ყველაზე სასარგებლო იყო ფილოგეოგრაფიულ კვლევაში, იმის გამო დამახასიათებელია, რომ ეს გენები არ არის რეკომბინირებული და შედეგად, ისინი აჩვენებენ ბევრად უფრო მკაფიო ფილოგენეტიკურ ხაზს, ვიდრე ბევრს. ბირთვული გენები. პოპულაციის ეფექტური ზომა, რომელიც გამოითვლება მიტოქონდრიული გენებით (და Y ქრომოსომა ანალოგიურად ფუნქციონირებს) მიტოქონდრიული გენების გამოთვლის დაახლოებით მეოთხედია. ბირთვული გენები, მაშინ დივერგენცია ხდება თითქმის ოთხჯერ უფრო სწრაფად, ვიდრე ბირთვული გენების შემთხვევაში, განსხვავების ამ სწრაფმა სიჩქარემ (და გენის ნაკადმა) შეიძლება გამოიწვიოს დაკვირვებული მემკვიდრეობის ნიმუში ამ ცალმხრივ გენებში განსხვავდება ბირთვული გენებით მიღებული ფილოგენიებისგან (რომლებიც წარმოადგენენ გენოფონდის უმრავლესობას ინდივიდუალური).
კავალი-სფორცა, ლ. ლ. (2000). გენები, ხალხები და ენები (1st Edition in Pocket Library ed.). ბარსელონა, ესპანეთი: კრიტიკული რედაქცია.
Rebato, E., Susanne, C., & Chiarelli, B. (რედაქტირება.). (2005). ბიოლოგიური ანთროპოლოგიის გაგება: ევოლუცია და ადამიანის ბიოლოგია. ღვთაებრივი სიტყვა.
ტორონი, ა., შური, თ. გ., კებელი, მ. ფ, ბრაუნი, მ. დ, ნილი, ჯ. ვ., ლარსენი, მ., და სხვ. (1993). აზიური აფინურები და კონტინენტური გამოსხივება ძირძველი ამერიკის ოთხი დამფუძნებელი mtDNA. The American Journal of Human Genetics, 53 (3), 563.
(2022). მიტოქონდრიული დნმ, ადამიანის გენომის ეროვნული კვლევითი ინსტიტუტი.
-მიტოქონდრიული დნმ-ის სქემატური მითითება-
კრესპილო, მ., პარედესი, მ., არიმანი, ჯ., გერერო, ლ., და ვალვერდე, ჯ. (2004). ესპანეთის სამოქალაქო ომი (1936-1939): კატალონიის მასობრივი საფლავებიდან ადამიანის ნაშთების იდენტიფიცირება მიტოქონდრიული დნმ-ის ანალიზით. საქმის შესახებ. სასამართლო მედიცინის რვეულები (38), 37-46.
