
08/08/2023

Mitohondrijski genom je majhna, zaprta krožna molekula z dvojno vijačnico s 16.569 baznimi pari (bp), ki se deduje le od matere na sinove in hčere (redko). primeri heteroplazme), zato v času spočetja ni genetske rekombinacije, razvija se tudi izključno s kopičenjem mutacij v vreme.





Visoka stopnja mutacije mitohondrijskega genoma (10- do 20-krat večja kot pri DNK jedrski, v smislu genov s primerljivimi funkcijami) je uporaben za razlikovanje med populacijami, ki se skozi čas so bili biološko povezani, vendar je njihova stopnja mutacije lahko tako hitra, da lahko pride do pojava mutacije. povratna mutacija. Stopnja evolucija Srednja vrednost te molekule je bila izračunana iz vrst, pri katerih so bili na voljo divergenčni časi iz fosilnih ostankov, in iz biodemografskih ali beljakovinskih podatkov; rezultat daje 1-2 % na milijon let, kar velja za različne rede.
Ta posebna DNK deluje kot orodje za raziskovanje
biološko sorodnost med populacijami, ker njihove lastnosti omogočajo razjasnitev odnosov med populacijami ki so se v zadnjem času razšli, ne da bi morali upoštevati pojave rekombinacije in dodati dimenzijo začasno; Za to je treba upoštevati naslednje: dva posameznika, katerih skupni prednik je bila ženska, bosta imela molekule mitohondrijske DNA tako različne, kolikor je pretekel čas od ločitve prednik.Nekatere regije DNK so lahko med seboj zelo podobne, kar omogoča njihovo razvrstitev v istem grozdu, ki je znan kot haploskupina. Torroni in drugi (1993) so na primer tipizirali ustanovne mitohondrijske linije za ameriško celino, ki so prejeli poimenovanje haploskupin A, B, C in D, glede na točkovne mutacije zaporedja mitohondrijski. Te mutacije ustvarijo različna mesta cepitve za specifične encime, kot je opisano spodaj: v liniji A je mesto meja 663 za encim HaeIII, linija C je označena z mestom 13259 za HincII, v liniji D je mesto 5176 prepoznano za AluI; ta identifikacija je narejena iz polimorfizmov dolžine fragmentov (RFLP). V primeru linije B je na položaju 8272-8289 izbrisanih 9 baznih parov.
Geografska porazdelitev vsake linije je bila opisana takole: linija A je prevladujoča na ameriški celini in zlasti v Severni Ameriki; vendar se ta haploskupina pripisuje tudi mezoameriškim populacijam. Liniji C in D se pojavljata predvsem v Južni Ameriki; linijo B najdemo v severni in južni regiji pacifiške obale in Kemp et al (2010) jo predlagajo kot značilno linijo družina Yuto-Aztecan in jugozahodne ZDA.
Poleg tega obstajajo mutacije, ki si jih delijo posamezniki iste haploskupine, kar je omogočilo opis specifičnih haplotipov (ali podlinij) nekaterih populacij. Slika 1 prikazuje področja mitohondrijske DNK, ki vsebujejo mutacije za prepoznavanje vsake linije.
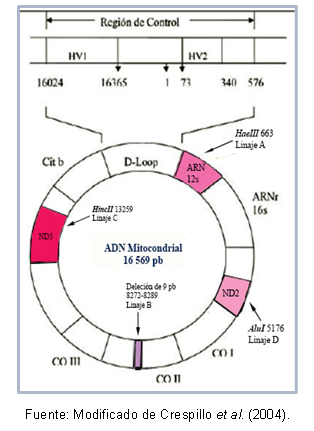
Slika 1. Regije mitohondrijske DNK, ki predstavljajo vsako ameriško ustanovno linijo.
Nekaj let so bile priznane samo te štiri linije, dokler ni bila vključena haploskupina X za severne populacije celine; ki je tudi v daljnem sorodstvu z evropskimi populacijami. Prejšnje linije se ujemajo z azijskimi populacijami, čeprav so opažene manj pogosto kot na ameriški celini; linij A, B in C ne najdemo pri sodobnih Afričanih in Kavkazijcih; in linija D obstaja tudi v Afriki, vendar povezana z drugimi restrikcijskimi mesti. Glede na navedeno bi lahko rekli, da so te haploskupine značilne za ameriške populacije in je zato njihova študija ustrezna z vidika poselitve in migracij (starih in nedavnih).
Način ugotavljanja genetskih podobnosti ali razlik med populacijami je z genetskimi razdaljami, ki imajo lahko a zgodovinsko razlago, saj se skozi generacije spreminjajo (večajo ali zmanjšujejo) in nas lahko pripeljejo do dogodkov v zgodovini enega prebivalstvo na primer: v času velike selitve ali stika med dvema kulturama lahko s tem izračunom razberemo, kateri mehanizmi so se aktivirali, da so nam dali ta ali oni rezultat.
Tako genetski drift kot genski tok sta povezana z lahkoto, s katero se mora skupina, v tem primeru človeška bitja, premikati znotraj skupine. ozemlje in stopiti v stik z drugimi skupinami. Geografska izolacija je torej genetska razdalja, ki raste z naraščajočo geografsko razdaljo med človeškimi skupinami.
Za razlago raznovrstnost v ustanovnih mitohondrijskih linijah v Ameriki sta dva hipoteza: da je bila ta celina kolonizirana zaradi številnih dogodkov iz Beringije ali da so se po kolonizaciji zgodile evolucijske spremembe, ko je prišlo do selitve. Pojasnjeni sta tudi dve poti vstopa teh variacij na celino, prva predlaga, da so štiri ustanovne haploskupine brez variacij, to je vsaka s korenskega haplotipa bi lahko prispeli tik po zadnjem ledeniškem maksimumu ali malo pred njim, z datumi pred 21 tisoč do 19 tisoč leti in bi sledili obalni poti mimo Mirno; drugi predlog nakazuje, da so te variacije znotraj haploskupine že obstajale v Beringiji in so bile prinesene na jug ameriške celine, vendar bi bil njihov vstop točno na koncu zadnjega ledeniškega maksimuma, tako da bi bile poti znotraj celine že proste, potem bi bil vstop teh človeških skupin pred 19 tisoč leta. Obstaja tudi velika raznolikost haploskupine A in njen koalescenčni čas, ki je krajši od ostalih (17 tisoč let).
Najverjetnejša razlaga za to je, da je to posledica sekundarnih širitev haploskupine A iz Beringije, dolgo po koncu zadnjega ledeniškega maksimuma. Kljub neskladju glede časa vstopa človeka v Ameriko so genetske študije dajejo nekaj jasnosti, ker podpirajo hipotezo, da so na ameriški celini že prej obstajale človeške skupine Clovis; in ugotovljeno je, da obstaja ločitev med predniki severovzhodne Azije pred 25-35 tisoč leti in vstopom v Ameriko pred 15-35 tisoč leti. Slika 2 prikazuje poti mitohondrijskih haploskupin v svetu in čas razhajanja v letih pred sedanjostjo.
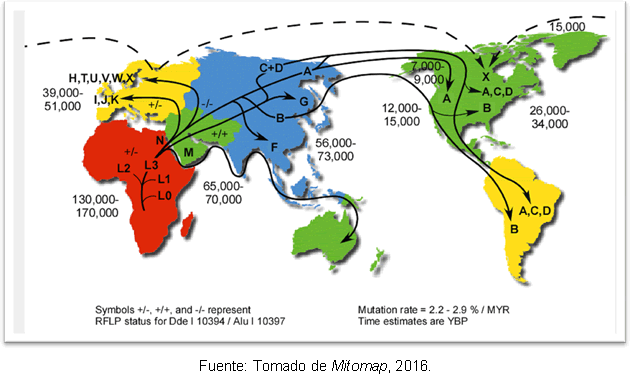
Slika 2. Zemljevid različnih poti širjenja mitohondrijskih rodov
Obstaja zelo uporabno orodje za genetsko analizo; filogeografija. To so prve aplikacije molekularnih študij in s tem poskušamo določiti filogenetska in prostorska razmerja med nukleotidnimi zaporedji, v tem primeru DNK mitohondrijski. Prostorska porazdelitev je lahko podobna časovnemu vzorcu, kar pomeni, da bi morala biti geografsko najbolj oddaljena zaporedja DNK najbolj genetsko različni, pa tudi zaporedja DNK, ki so se že zdavnaj ločila, bi morala biti tudi najbolj različna genetsko. Tako bi geografsko oddaljene populacije z malo ali nič pretoka genov med njimi kopičile razlike zaradi genetskega premika in mutacije, tudi s selekcijo; lahko pa pride do situacij, ki ne dopuščajo premikanja, kot je eden ali več učinkov ustanovitelja ali drugih vzorcev pretoka genov.
Podatki, ki jih zagotavljajo mitohondrijski geni, so bili med najbolj uporabnimi v filogeografskih raziskavah zaradi značilnost teh genov je, da se ne rekombinirajo in posledično kažejo veliko jasnejšo filogenetsko linijo kot mnogi jedrski geni. Dejanska velikost populacije, izračunana z mitohondrijskimi geni (in s kromosomom Y, ki deluje na podoben način), je približno četrtina tiste, izračunane za mitohondrijske gene. jedrskih genov, potem pride do divergence skoraj štirikrat hitreje kot pri jedrskih genih, lahko ta hitra stopnja divergence (in pretoka genov) povzroči Opaženi vzorec dedovanja v teh enostarševskih genih je drugačen od filogenije, pridobljene z jedrnimi geni (ki predstavljajo večino genskega sklada v posameznik).
Cavalli-Sforza, L. L. (2000). Geni, ljudstva in jeziki (1. izdaja v izdaji Pocket Library). Barcelona, Španija: kritični uvodnik.
Rebato, E., Susanne, C. in Chiarelli, B. (Urejanje.). (2005). Razumeti biološko antropologijo: evolucijo in človeško biologijo. Božanska beseda.
Torroni, A., Schurr, T. G., Cabell, M. F, Brown, M. D, Neel, J. V., Larsen, M., et al. (1993). Azijske afinitete in celinsko sevanje štirih ustanovnih indijanskih mtDNA. American Journal of Human Genetics, 53(3), 563.
(2022). mitohondrijska DNK, Nacionalni raziskovalni inštitut za človeški genom.
-Referenca sheme mitohondrijske DNK-
Crespillo, M., Paredes, M., Arimany, J., Guerrero, L., & Valverde, J. (2004). Španska državljanska vojna (1936-1939): identifikacija človeških posmrtnih ostankov iz množičnih grobišč v Kataloniji z analizo mitohondrijske DNK. O primeru. Sodnomedicinski zvezki (38), 37-46.

