01/02/2022
0
ビュー

ミトコンドリアゲノムは、16,569塩基対(bp)の小さな閉じた二重らせん環状分子であり、母親から息子と娘にのみ継承されます(まれに)。 ヘテロプラスミーの場合)、受胎時に遺伝子組換えがないため、突然変異の蓄積によってのみ進化します 天気。





ミトコンドリアゲノムの高い変異率(10〜20倍 DNA 核は、同等の機能を持つ遺伝子の観点から)、時間の経過とともに集団を区別するのに役立ちます 生物学的に関連していますが、それらの突然変異率は非常に速いため、突然変異現象が発生する可能性があります。 バックミューテーション。 の割合 進化 この分子の平均は、化石の残骸からの発散時間が利用可能な種から、および生物人口統計学的またはタンパク質データから計算されています。 その結果、100万年あたり1〜2%が得られ、さまざまな注文に有効です。
この特定のDNAは、 リサーチ それらの特性が集団間の関係を解明することを可能にするので、集団間の生物学的親族関係 組換え現象を考慮したり、次元を追加したりすることなく、最近分岐したもの 一時的; このため、次の点に注意する必要があります。共通の祖先が女性である2人の個人が ミトコンドリアDNA分子は、分離してから時間の経過とともに変化します。 祖先。
DNAのいくつかの領域は互いに非常に類似している可能性があり、 分類 同じクラスター内で、ハプログループとして知られています。 たとえば、Torroni et al(1993)は、アメリカ大陸のミトコンドリア系統の創設を代表しました。 シーケンスの点突然変異に応じて、ハプログループA、B、C、およびDの宗派を受け取りました ミトコンドリア。 これらの変異は、以下に説明するように、特定の酵素に対して異なる切断部位を作成します。系統Aには部位があります 酵素HaeIIIのカットオフ663、系統CはHincIIのサイト13259によって特徴付けられ、系統Dではサイト5176が認識されます AluI; この識別は、フラグメント長多型(RFLP)から行われます。 系統Bの場合、8272-8289の位置で9塩基対の欠失があります。
各系統の地理的分布は次のように説明されています。系統Aはアメリカ大陸、特に北アメリカで優勢です。 ただし、このハプログループはメソアメリカの人口にも起因しています。 系統CとDは主に南アメリカに出現します。 系統Bは太平洋岸の北部と南部に見られ、Kemp et al(2010)はそれを
家族 ユト・アステカンとアメリカ南西部さらに、同じハプログループの個人間で共有される突然変異があり、これにより、一部の集団の特定のハプロタイプ(または亜系統)を説明することが可能になりました。 図1は、各系統を認識するための変異を含むミトコンドリアDNAの領域を示しています。
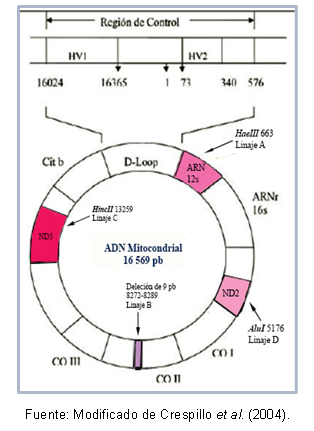
図1。 各アメリカの創設系統を代表するミトコンドリアDNA領域。
大陸の北部の個体群にハプログループXが含まれるまで、数年間はこれら4つの系統のみが認識されていました。 これもヨーロッパの人口とは遠い関係にあります。 以前の系統はアジアの個体群と対応していますが、アメリカ大陸ほど頻繁には観察されていません。 系統A、B、およびCは、現代のアフリカ人および白人には見られません。 系統Dもアフリカに存在しますが、他の制限部位に関連付けられています。 以上のことから、これらのハプログループはアメリカの人口を特徴づけるものであり、したがって彼らの研究は定住と移住(古いものと最近のもの)の観点から適切であると言えます。
集団間の遺伝的類似性または相違性を確立する方法は、遺伝的距離を使用することです。 それらは世代を超えて変化(増加または減少)し、歴史上の出来事につながる可能性があるため、歴史的説明 ひとつの 人口 たとえば、2つの文化間の大規模な移行または接触のときに、この計算を使用して、これまたはその結果をもたらすためにどのメカニズムが実行されたかを識別できます。
遺伝的浮動と遺伝子流動はどちらも、グループ(この場合は人間)がグループ内を移動しなければならないことの容易さに関係しています。 地域 そして他のグループと接触すること。 したがって、地理的隔離は、人間のグループ間の地理的距離の増加に伴って増加する遺伝距離です。
説明するには 多様性 アメリカでのミトコンドリア系統の創設には、2つあります 仮説:この大陸はベーリング地峡からの複数の出来事によって植民地化されていること、または移住が発生すると、植民地化後に進化の変化が発生したこと。 これらのバリエーションが大陸に入る2つのルートも説明されています。最初のルートでは、バリエーションのない4つの創設ハプログループ、つまり、それぞれにバリエーションのあるハプログループがあります。 ルートハプロタイプ、最終氷期最盛期の直後または少し前に到着した可能性があり、日付は21,000〜19000年前であり、沿岸ルートをたどっていたはずです。 平和的; 2番目の提案は、これらのハプログループ内のバリエーションがすでにベーリング地峡に存在し、アメリカ大陸の南に持ち込まれたことを示唆していますが、それらのエントリは 最終氷期最盛期の終わりに、大陸内のルートがすでに無料になるようにすると、これらの人間グループの侵入は1万9千年前になります。 年。 また、ハプログループAには非常に多様性があり、他のハプログループよりも短い合体時間(17000年)があります。
これについての最も可能性の高い説明は、最終氷期最盛期の終了後ずっと、ベーリング地峡からのハプログループAの二次拡大によるものであるということです。 人間が南北アメリカに入る時期に関する不一致にもかかわらず、遺伝学の研究は 彼らは以前にアメリカ大陸に人間のグループがあったという仮説を支持しているので、ある程度の明確さを与えてください クロービス; そして、25-35千年前の北東アジアの祖先と15-35千年前のアメリカへの入国との間には隔たりがあることがわかります。 図2は、世界のミトコンドリアハプログループの経路と、現在より数年前の分岐の時期を示しています。
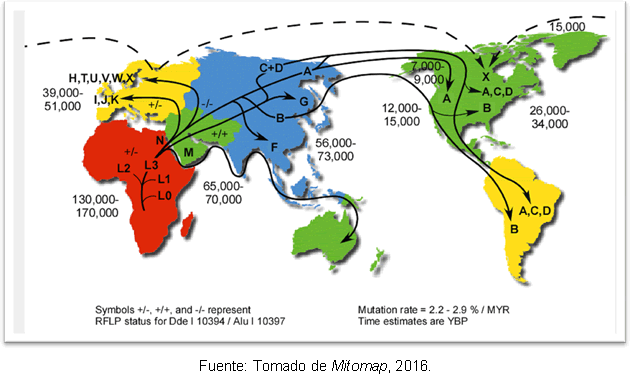
図2。 ミトコンドリア系統の分散のさまざまなルートのマップ
遺伝子解析には非常に便利なツールがあります。 系統地理学。 これらは分子研究の最初のアプリケーションであり、それを使用して決定できるように努めています ヌクレオチド配列、この場合はDNA間の系統発生的および空間的関係 ミトコンドリア。 空間分布は時間的パターンに似ている可能性があります。つまり、地理的に最も離れたDNA配列が最も多いはずです。 遺伝的に異なるだけでなく、はるか昔に分岐したDNA配列も最も異なるはずです 遺伝的に。 したがって、地理的に離れた集団は、それらの間に遺伝子流動がほとんどまたはまったくないため、選択によってさえ、遺伝的浮動および突然変異による差異を蓄積します。 ただし、1つ以上の創始者効果や他の遺伝子流動パターンなど、ドリフトを許可しない状況が発生する場合があります。
ミトコンドリア遺伝子によって提供されるデータは、系統地理学の研究で最も有用なものの1つです。 これらの遺伝子は再結合しないという特徴があり、その結果、多くの遺伝子よりもはるかに明確な系統発生線を示します 核遺伝子。 ミトコンドリア遺伝子で計算された(そしてY染色体が同様に機能している)有効個体数は、ミトコンドリア遺伝子で計算されたものの約4分の1です。 核遺伝子の場合、発散は核遺伝子の場合よりもほぼ4倍速く発生します。この急速な発散(および遺伝子流動)により、 これらの片親性遺伝子で観察された遺伝形式は、核遺伝子で得られた系統発生とは異なります(核遺伝子( 個人)。
カヴァッリスフォルツァ、L。 L。 (2000). 遺伝子、人々、言語(Pocket Library ed。の第1版)。 バルセロナ、スペイン:重要な社説。
Rebato、E.、Susanne、C.、およびChiarelli、B. (編集)。 (2005). 自然人類学を理解するために:進化と人間生物学。 神の言葉。
Torroni、A.、Schurr、T. G.、Cabell、M。 F、ブラウン、M。 D、ニール、J。 V.、Larsen、M.、etal。 (1993). 4つの創設ネイティブアメリカンmtDNAのアジアの親和性と大陸の放射線. American Journal of Human Genetics、53(3)、563。
(2022). ミトコンドリアDNA、国立ヒトゲノム研究所。
-ミトコンドリアDNAスケマティックリファレンス-
Crespillo、M.、Paredes、M.、Arimany、J.、Guerrero、L.、およびValverde、J. (2004). スペイン内戦(1936-1939):ミトコンドリアDNA分析によるカタルーニャの集団墓地からの人間の遺体の識別。 ケースについて. 法医学ノートブック(38)、37-46。
